The Sima de las Palomas vertical cave system was formed by karst solution of a Triassic marble hill in present-day Spain overlooking the Mediterranean. In 1991, a speleologist descending the 18-m deep entrance shaft (Fig.1) extracted a fossil (designated SP1) which, once the cemented breccia was removed, comprised Neanderthal maxillae connected to the mandible, with almost all their adult teeth. Subsequent systematic excavation (Walker et al., 2012a) uncovered three undisturbed Neanderthal partial skeletons (SP96, SP92, and a child, SP97) with several skeletal parts in anatomical position (including cranio-mandibular articulation, femoro-pelvic articulation, elbow, rib-cage, vertebral column, shoulder girdle, foot bones, etc.), Mousterian Palaeolithic artifacts, and animal bones (some charred), all lying deeply in a cemented rock tumble within the upper part of an 18-m deep wall of brecciated sediments (Fig. 2) was left exposed by miners after they took out most of the shaft’s sedimentary fill ca. 1900. SP96 and SP97 have crania and mandibles (Figs. 3, 4), unlike SP92. Because SP1 had lain near SP92 it might be SP92’s head. Excavation uncovered SP96 lying with elbows flexed and hands touching the forehead. Computer-assisted tomography revealed hand-bones in breccia adhering to the forehead of the SP97, identified as a child (Walker et al., 2012b), which lay underneath SP96, an adult, perhaps its parent. The position of the upper extremities implies intentional arrangement before rigor mortis had set in; it is recorded at some other Mousterian sites (Defleur, 1993).
______________________________________
 Fig. 1: Sima de las Palomas del Cabezo Gordo: Main shaft.
Fig. 1: Sima de las Palomas del Cabezo Gordo: Main shaft.
_____________________________________
 Fig. 2: Excavation area A: Mouth of Main Shaft. B: Excavation area C: Base of rock tumble that contained the three Neanderthal skeletons. D: Thin hard conglomerate band beneath which excavation is continuing in layers with Mousterian remains.
Fig. 2: Excavation area A: Mouth of Main Shaft. B: Excavation area C: Base of rock tumble that contained the three Neanderthal skeletons. D: Thin hard conglomerate band beneath which excavation is continuing in layers with Mousterian remains.
_____________________________________
 Fig. 3: Neanderthal adult female skeleton SP96, “Paloma”
Fig. 3: Neanderthal adult female skeleton SP96, “Paloma”
____________________________________
 Fig. 4: Neanderthal child skull SP97 (“Paloma’s child”)
Fig. 4: Neanderthal child skull SP97 (“Paloma’s child”)
_____________________________________
Individual articulated skeletons are very important indeed because they provide more precise and accurate estimates of body size and proportions than estimates derived from pooled statistical analyses of a given bone type (e.g. femur, tibia, or humerus) from an assemblage of bones from different people who may well not be identifiable individually. SP96 was a short Neanderthal woman (”Paloma”), less than 20 years old, with the typically female wide greater sciatic notch of the pelvic basin which underwent post mortem distortion (that “virtual” reconstruction will correct from tomographs). The skeleton is about 85% complete, which permits precise and accurate morphological measurements for estimating body proportions (e.g. crural index, etc.) which are undoubtedly Neanderthal and robust (“hyperpolar”). Remarkably, its stature is one of the shortest known for Neanderthal adults (Walker et al., 2011a).
The child skeleton, SP97, lay underneath “Paloma” (her child?). Also short in stature, SP92 was probably less than 25 years old (Walker et al., 2011b). Short, too, was the owner of SP77, a femoral head from looser sediment that had banked up against cemented rock tumble and contained scattered Neanderthal remains, including mandibular fragments of a baby, a child, and an adolescent female (Walker et al., 2010a; Walker et al., 2008); this sediment contained lenses with signs of burning. Fragments of three more Neanderthal mandibles were found while sieving rubble left by miners. The Neanderthal finds included many teeth and bone fragments.
In all, at least 9 Neanderthals are represented at the site. The excavated Neanderthal remains correspond to a time about 50,000 years ago; different scientific methods give estimates implying >40,000-<60,000 (U-series, TL, 14C: for details, see Walker et al., 2012b). Palaeopalynology indicates cool moist conditions though with persistence of species ill-adapted to resist frost (Carrion et al., 2003).
______________________________________
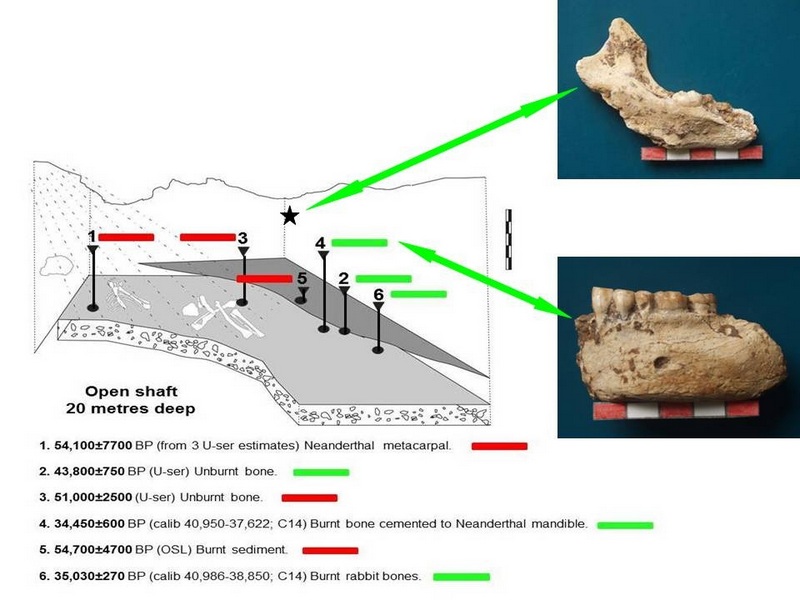 Fig. 5: Stratigraphical position of the three articulated Neanderthal skeletons and situations of dated materials and two other Neanderthal mandibles (for further information, see Walker et al., 2012a).
Fig. 5: Stratigraphical position of the three articulated Neanderthal skeletons and situations of dated materials and two other Neanderthal mandibles (for further information, see Walker et al., 2012a).
_______________________________________
Excavated with the SP97 child were the only articulated bones of large animals found at the site so far, viz., two sets of horse ankles (calcaneum, talus, cuboid), one group, cemented by CaCO3 to SP97’s skull, had undergone burning, the other, unburnt, beneath SP97’s trunk, included additionally a distal tibial fragment, and a third horse talus lay underneath SP97. Two leopard metacarpal bones lay near SP97’s skull and two leopard hind-paws, with metatarsal and phalangeal bones in anatomical articulation, lay 0.5 m from SP97’s skull in a similar level. It is unlikely horse ankles and leopard paws were fortuitous accumulations, given absence around SP97 of other body parts of those species. A heavily burnt leopard temporal bone found in mine rubble implies Neanderthal intervention; a large premolar tooth implies presence of cave hyaenas. Flakes and spalls from flint-knapping lay close to SP97. The site has yielded some carefully prepared Levallois points on flat triangular flint flakes with finely-retouched margins, surely effective for hunting with thrusting spears. Flint outcrops are unknown on Cabezo Gordo. Some flint quite likely came from a hydrothermal outcrop inspected ca. 25 km to the south, though it is hardly the source of the bulk of the Palaeolithic assemblage. Despite lack of clear-cut signs of intentional interment, large stones might have been thrown over SP96, SP92 and SP97 to deter leopards and hyaenas from disturbing the corpses. This is a prosaic interpretation.
Rarely are Neanderthal skeletons uncovered in anatomical connection. When animals die their soft parts decompose quickly, aided and abetted by various organisms (carnivorous animals or birds, insects, saprophytic fungi, bacteria, etc.), after which skeletons come apart. Long before they can get buried by natural deposition of sediment, wind and rain may scatter bones where carnivorous animals and birds have failed (exceptionally, skeletons of creatures trapped in caves or swamps escape from being scattered). At Sima de las Palomas three articulated skeletons from 50,000 years ago lay close together. It raises a conjecture that behavioural or cultural impingement occurred, implicating individuals other than the three deceased. Maybe it is an instance of Neanderthals attending to their dead, albeit with a prosaic motive.
The three skeletons and the rock tumble over them lay on a thin bed of extraordinarily hard conglomerate, though it contained some Palaeolithic artifacts and charred bone fragments. Beneath it coarse sediment has been excavated to a depth of 2 m so far, containing many bones of red deer, horse and other herbivores, many of which are charred, as well as rabbits and other small animals, including mandibular fragments of two porcupines (Hystrix brachyura: Rhodes et al., 2013).Tortoise seems to have played a part in the diet (Morales-Pérez and Sanchis-Serra, 2009). Characteristically Mousterian flint artifacts are present. Several smooth rounded cobbles were undoubtedly brought by Neanderthals to the site from stream gravels in the plain below. Being larger than some hammer-stones from the site they might have been used to pound or grind minerals (perhaps haematite; the Cabezo Gordo marble contains veins of magnetite and other iron ores) or foodstuff. Vegetable food at Sima de las Palomas is suggested both by phytoliths discovered in calculus on some Neanderthal teeth (Salazar-García et al., 2013) and two examples of dental caries (Walker et al., 2010b).
________________________________________
Acknowledgements:
We thank all our research collaborators named in the text or references, but above all Dr. E. Trinkaus for his tireless work. Among our many other collaborators we thank archaeologists Dr. D.A. Roe and I. Martín Lerma, palaeontologist Dr. X. Murélaga Bereicua, and anthracologist Dr. E. Badal García.
Legends to figures:
Fig. 1 Sima de las Palomas del Cabezo Gordo: Main shaft.
Fig. 2 Sima de las Palomas del Cabezo Gordo: Excavation area. A Mouth of Main Shaft. B Excavation area. C Base of rock tumble that contained the three Neanderthal skeletons. D Thin hard conglomerate band beneath which excavation is continuing in layers with Mousterian remains.
Fig. 3 Sima de las Palomas del Cabezo Gordo: Neanderthal adult female skeleton SP96, “Paloma”. SP96 has parts of all skeletal units except feet. Open L (left) greater sciatic notch shows SP96 was female. Full occlusal eruption of L M3 and ventrally unfused sacral bodies, unfused proximal clavicular epiphysis, and partially fused iliac crest, imply death at ≤20 years. All sufficiently-preserved long-bone and manual epiphyses are fused. SP96 skull had undergone crushing with loss of mandibular corpus and posterior, inferior and most of L neurocranium. R (right) facial elements are undistorted, albeit broken and displaced, with 22 teeth cemented into them. Axial skeleton comprises partial lower cervical, upper thoracic, and lumbar vertebrae; 5 sacral bodies; and >6 R ribs cemented to R scapula and clavicle which are complete, as are all 6 upper limb long bones. Metacarpals and manual phalanges are mostly present in anatomical position (some carpals show erosion). Posterior ilum apart, all elements of ossa coxae are present on at least one side, albeit broken, displaced, and cemented. R and L femora and L tibia are almost complete. Only SP96 and La Ferrassie 1 afford reliable adult Neanderthal lengths of all 4 major limb segments, clavicular and scapular lengths, and weight-bearing articular dimensions. SP96 Neanderthal cranial features: lateral supraorbital torus; broad supratoral sulcus; large frontozygomatic suture with columnar frontal process of zygoma; little horizontal angulation of anterolateral zygoma; absent canine fossa; strongly bilevel nasal floor with sharply-angled inferior nasal aperture margin; nasal aperture breadth (30–32 mm) overlapping between Neanderthal and Upper Palaeolithic modern human values; mandible with high R coronoid process; mandibular notch with lowest point closer to condyle; prominent superior medial pterygoid tubercle on ramus; rounded L gonial angle; strongly shovelled I2 with marked labial convexity. Neanderthal postcranial features: strong dorsal sulcus of the right scapula; narrow medial humeral pillar; medially orientated radial tuberosity; subequal pollical phalangeal lengths; ulnar deviation of pollical distal phalanx; elongated superior pubic ramus with thinned ventral margin; absent femoral pilasteric development; rounded convex cross-section of tibial diaphysis; absence of distinct fibular diaphyseal sulci. Aspects of SP96 less common in Neanderthals: nontaurodont teeth; short I1 labial root length (SP19 and SP21 have Neanderthal values); absent opponens pollicis flange on metacarpal 1; manual terminal phalanges of digits 3 and 4 with apical tufts lacking the Neanderthal rounded curve (e.g. SP28) and distinct ungual spines.
Fig. 4 Sima de las Palomas del Cabezo Gordo: Neanderthal child skull SP97 (“Paloma’s child”).
Fig. 5 Sima de las Palomas del Cabezo Gordo: Stratigraphical position of the three articulated Neanderthal skeletons and situations of dated materials and two other Neanderthal mandibles (for further information, see Walker et al., 2012a).
Fig. 6 Sima de las Palomas del Cabezo Gordo: Exploded stratigraphical sections and site plan; the thin hard conglomerate band in Fig. 2D runs across the middle of the sections.
References:
Carrión, J.S., Yll, E.I., Walker, M.J., Legaz, A.J., Chain, C., López, A., 2003. Glacial refugia of temperate, Mediterranean and Ibero-North African flora in south-eastern Spain: new evidence from cave pollen at two Neanderthal man sites. Global Ecology and Biogeography12: 119-129.
Defleur, A., 1993. Les sépultures moustériennes, Paris, Éditions du Centre National de la Recherche Scientifique.
Morales-Pérez, J.V. and Sanchis-Serra, A., 2009. The Quaternary fossil record of the genus Testudo in the Iberian Peninsula. Archaeological implications and diachronic distribution in the western Mediterranean. Journal of Archaeological Science31: 1152-1162.
Rhodes, S.E., Walker, M.J., López-Martínez, M., Haber-Uriarte, M., López.Jiménez, A., Buitrago-López, A.T., Dewar, G., 2013. Analysis of Hystrix specimens recovered from Sima de las Palomas, Murcia, Spain, in Program with Abstracts, Canadian Association for Physical Anthropology, 41st Annual Meeting, October 17-20 2013 Scarborough, ON (University of Toronto, Scarborough).Scarborough, University of Toronto, and Canadian Association for Physical Anthropology, p. 47.
Salazar-García, D.C., Power, R.C., Sanchis Serra, A., Villaverde, V., Walker, M.J., Henry, A.G., 2013. Neanderthal diets in central and southeastern Mediterranean Iberia. Quaternary International318,3-18.
Walker, M.J., López Martínez, M., Ortega-Rodrigáñez, J., Haber-Uriarte, M., López-Jiménez, A., Avilés-Fernández, A., Polo-Camacho, J.L., Campillo-Boj, M., García-Torres, J., Carrión-García, J.S., San Nicolas-del Toro, M., Rodríguez-Estrella, T., 2012a. The excavation of the buried articulated Neanderthal skeletons at Sima de las Palomas (Murcia, SE Spain). Quaternary International 259: 7-21.
Walker, M., Ortega Rodrigáñez, J., Agut Giménez, A., Soler Laguía, M., Zollikofer, C.P.E., Ponce de León, M.S. 2012b. The Sima de las Palomas Neanderthal skeletons: First steps towards “virtual” reconstruction. Proceedings of the European Society for the Study of Human Evolution 1: 191 (special issue, J-J.Hublin, W.Roebroeks, M.Soressi, T.Terberger, F.Spoor, Eds., Proceedings of the 2nd Annual Meeting of the European Society for the Study of Human Evolution ESHE 21-22 September 2012 Bordeaux/France). Leipzig, Max-Planck Institute for Evolutionary Anthropology, Society for the Study of Human Evolution.
Walker, M.J., Ortega, J., Parmová, K., López, M., Trinkaus, E., 2011a. Morphology, body proportions, and postcranial hypertrophy of a female Neandertal from the Sima de las Palomas, southeastern Spain. Proceedings of the National Academy of Sciences USA 108:10087–10091.
Walker, M.J., Ortega Rodrigáñez, J., López Martínez, M., Parmová, K., Trinkaus, E., 2011b. Neandertal postcranial remains from the Sima de las Palomas del Cabezo Gordo, Murcia, southeastern Spain. American Journal of Physical Anthropology 144: 505-515.
Walker, M.J., Lombardi, A.V., Zapata, J., Trinkaus, E., 2010a. Neandertal mandibles from the Sima de las Palomas del Cabezo Gordo, Murcia, southeastern Spain. American Journal of Physical Anthropology 142: 261-272.
Walker, M.J., Zapata, J., Lombardi, A.V., Trinkaus, E., 2010b. New evidence of dental pathology in 40,000 year old Neandertals. Journal of Dental Research90: 428-432.
Walker, M.J., Gibert, J., López Martínez, M., Lombardi, A.V., Pérez-Pérez, A., Zapata, J., Ortega, J., Higham, T., Pike, A., Schwenninger, J-L., Zilhão, J., Trinkaus, E., 2008. Late Neandertals in Southeastern Spain: Sima de las Palomas del Cabezo Gordo, Murcia, Spain. Proceedings of the National Academy of Sciences USA, 105: 20631-20636.
Michael Walker*†, Mariano López-Martínez**†, María Haber-Uriarte***†
© All rights reserved.
*Department of Zoology and Physical Anthropology, Biology Faculty, Murcia University, Campus Universitario de Espinardo Edificio 20, 30100 Murcia, Spain.
Email: [email protected] Tel: 34-620-267104
**Calle Pintor Joaquín 10-4º-I, 30009 Murcia, Spain.
Email: [email protected] Tel: 34-630-408806
***Department of Prehistory, Archaeology, Ancient History, Mediaeval History and Historiographical Techniques and Science, Faculty of Letters, Murcia University, Campus Universitario de La Merced, Calle Santo Cristo 1, 30001 Murcia, Spain.
Email: [email protected] Tel: 629-756183
†Directors of the excavation, Murcian Association for the Study of Palaeoanthropology and the Quaternary, MUPANTQUAT web-site http:www.mupantquat.com (Murcia Archaeological Museum, Avenida Alfonso X El Sabio 7, 30008 Murcia, Spain), all correspondence to MUPANTQUAT Secretary M.López Martínez <[email protected]>, and
†Murcia University Experimental Sciences Research Group E005-11 “Quaternary Palaeoecology, Palaeoanthropology and Technology” (c/o Dr.J.S.Carrión García, Department of Plant Biology, Biology Faculty, Murcia University, Campus Universitario de Espinardo Edificio 20, 30100 Murcia, Spain)
_____________________________________________________




